For organisms that live a life immersed in moving fluids, many biological functions such as respiration, photosynthesis and calcification, depend on the uptake of dissolved nutrients and/or gases. These uptake rates, in turn, are potentially limited by: 1) the transport rates of dissolved material from the water column to the surface of an organism (known as mass transfer limitation); 2) reaction kinetics at the boundary that limit the ability of an organism to assimilate the dissolved material across the body wall (reaction kinetic limitation), or 3) a combination of both. Using the barnacle, Balanus glandula, I explored the influence of water temperature and velocity on respiration to predict the environmental conditions under which barnacles are mass transfer limited.
Currently, I am investigating the effects of water motion on mass transfer in bilvalves and macrophytes. Techniques employed for these projects include particle image velocimetry and finite element modeling.
Movie 1.Does mussel feeding behavior increase local mixng? A 2D finite element model that pairs k-ε turbulence and convection-diffusion models to predict scalar transport based on the hydrodynamic characteristics measured for mussel exhalant jets.
Movie 2.Motions of individual Heterosigma cells recorded with a CCD video camera in a large tank that allowed for natural, free swimming behavior. Each track is 180 seconds from an individual cell of ~10 micron diameter. These algae swim in helical patterns (due to flagella), so there is a left-right waggle to their motion.
Movie 3. The induction of a surface aggregation of Heterosigma akashiwo depends on interaction between behavior and conditions in the hydrodynamic environment. The left beaker contains seawater and the right beaker contains a low-salinity layer over seawater. The brown coloration represents Heterosigma cells and the numbers indicate minutes. Cells on the right swim up to the surface, through the pycnocline and quickly become trapped in the low salinity surface layer. Cells on the left swim to the surface, aggregate and sink back down. You can observe continued mixing in the left beaker throughout the experiment, whereas the cells on the right become concentrated in a surface aggregation. Does swimming behavior represent a mechanism of algal bloom formation?
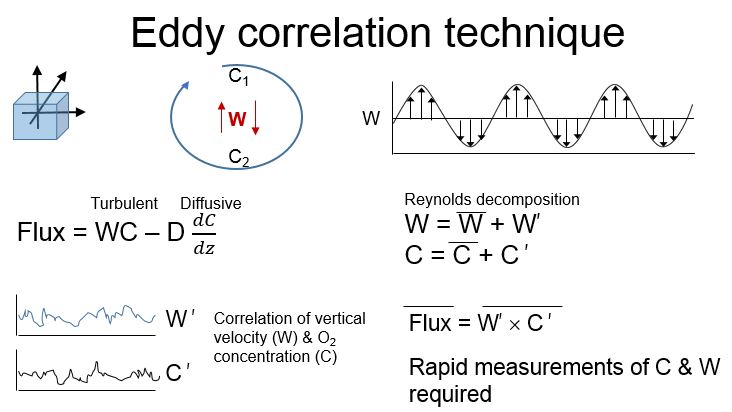
BENTHIC METABOLISM & EDDY CORRELATION
Anthropogenic nutrient loading has led to eutrophication of many aquatic ecosystems around the world. Although progress has been made in nutrient abatement from urban areas, cottage communities and agricultural lands, recovery in coastal ecosystems may follow a range of ecological trajectories. Internal loading alone may provide sufficient nutrients for persistent eutrophication, even when external loads have been controlled. This can lead to low hypolimnetic oxygen concentrations, endangering the health and survival of juvenile fish and benthic invertebrates. Thus, direct field measurements of biogeochemical flux are necessary to build effective hydrodynamic-biogeochemical models. Traditional methods of measuring benthic flux involve methods that disrupt natural hydrodynamic, biological and light conditions. Such limitations bias estimates of benthic flux, hindering predictions concerning ecosystem responses to changes in nutrient loading.
To determine whether internal nutrient fluxes counteract decreases in external loading, I employ a non-invasive eddy correlation technique (EC) to estimate O2 flux across the interface between benthic habitats and overlying water columns. The approach draws upon the statistical correlation between changes in scalar concentration (e.g., O2) and vertical velocities associated with turbulent eddies. EC estimates flux under natural hydrodynamics and light conditions, without interfering with natural benthic communities. Indeed, the EC device can measure flux above irregularly shaped mussel or macrophyte beds. The system can also be fitted to measure other scalars to estimate changes in parameters such as fluorescence or pH.